
Human Digestion: Main & Accessory Organs
In a previous article, I discussed the role of the enteric nervous system in regulation of muscle contraction of the human digestive system. Breaking digestive processes down by reviewing the role of each of its master control systems, makes the complexity of the digestive system easier to learn. In this article, I will consider the role of exocrine secretions to the digestive process and the human digestive system’s unique pattern of venous blood flow. In a following article, I will examine the role of endocrine hormones in the digestive process.
Human digestive system
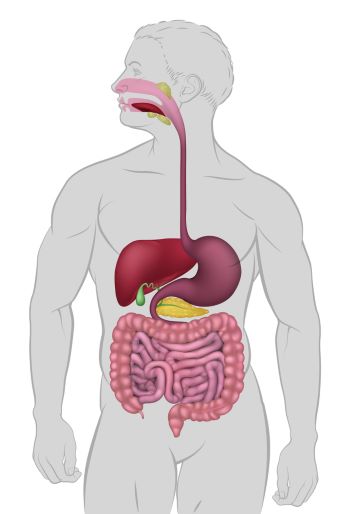
Human digestive organs, Christos Georghiou/Shutterstock.com
The digestive system’s main organs include the mouth, esophagus, stomach and intestines. Accessory digestive system organs are the pancreas, liver and gall bladder. Each of these organs depends upon the circulatory system to support its role in the digestion process.
There are six classes of nutrients absorbed by the gastrointestinal tract. They are water, fats, proteins, carbohydrates, vitamins and minerals. Each has its own pathway through the layers of the digestive tract and into the blood or lymph capillaries.
Breakdown of large food particles into molecules that can be transported by blood, requires secretion of digestive fluids by exocrine glands into the lumen of the gastrointestinal tract. The water source for secreted digestive fluids is the circulatory system’s capillaries.
Chewing and swallowing
The digestion process begins with chewing and swallowing at the mouth and esophagus. The muscles of the jaw, lips, tongue and cheek participate in chewing particles of food, which is called mastication. Coordination of the muscle activity to place food between the rows of teeth is accomplished by a group of neurons in the brain stem.
Chewing grinds food into smaller particles. It increases the surface area available to enzymes in saliva. Chewing also reduces the size of food particles so that they can be swallowed. The cellulose cell walls of vegetable matter are broken by pounding of the teeth, which is useful because humans cannot digest cellulose. And even more important to a continued desire to eat, chewing moves food about the mouth for sampling by taste receptors.
Saliva production
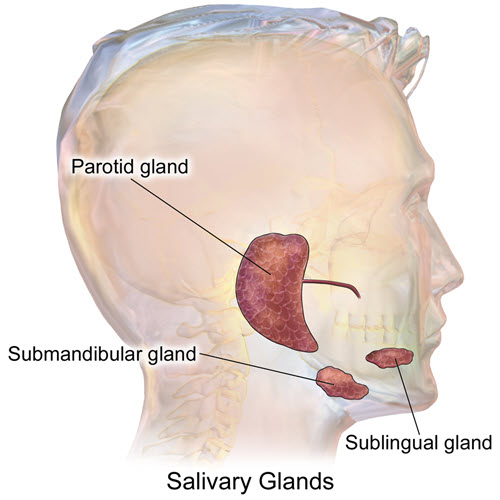
Human salivary glands, Bruce Blaus/Wikimedia Commons
Saliva is produced by the parotid glands near the back of the jaw, the sublingual glands and the submandibular glands. Sublingual glands lie under the tongue and the submandibular glands under the mandible.
Saliva moistens, lubricates and partially digests food particles. The parotid gland produces a watery, serous secretion that is about 50% of the saliva. Its serous secretion includes amylase, an enzyme that breaks down carbohydrate.
In humans, the sublingual and submandibular glands secrete a mixture of serous and mucus fluids to lubricate the food to ease swallowing.
How food particles are moved from the mouth to, and through the rest of the digestive tract by enteric neural activity can be found at ‘Gastrointestinal Tract Neural Regulation’.
Sensory afferents to the salivary nuclei of the brain stem originate at the taste buds, olfactory receptors and mechanoreceptors of the mouth. Activity of the salivary glands is increased by parasympathetic nerves of the autonomic nervous system originating in the brain stem. Remember that nerves are bundles of neuron axons and that nerves like those of the autonomic nervous system contain axons of both afferent and efferent neurons.
Chemicals released by parasympathetic nerves cause release of Ca++ from the endoplasmic reticulum of salivary gland cells. Increased intracellular Ca++ in turn stimulates a variety of ion transport systems that create an osmotic gradient to draw water from blood into the lumen of the glands.
Digestion in the stomach
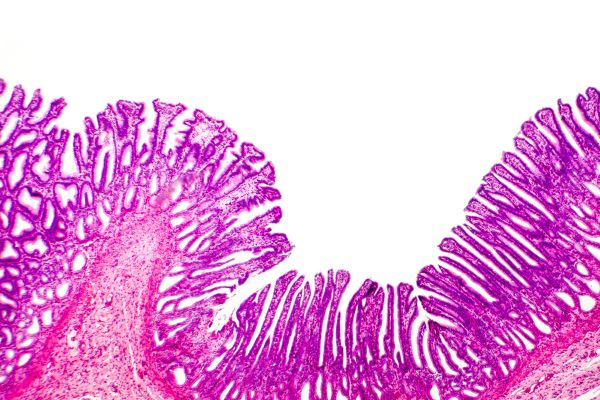
Histology of the gastric mucosa showing gastric pits, Kateryna Kon/Shutterstock.com
Digestion in the stomach begins with distension of the stomach wall by food intake. Contraction of the stomach muscles mixes and breaks apart food particles. Stomach secretions begin disruption the molecular bonds of proteins and lipids.
The lumen of the body of the stomach is lined by long glandular structures called gastric pits. Cells of the gastric pits secrete hydrochloric acid [HCl], the proteolytic enzyme precursor pepsinogen, mucus, gastric lipase and intrinsic factor.
Cells of the gastric pits closest to the lumen of the stomach secrete mucin and bicarbonate buffer. The mucin forms a gel that protects the cells of the stomach from the acid it secretes. Beneath the mucin, a layer of bicarbonate buffer is trapped to neutralize any [H+] penetrating the gel.
Gastric acid is secreted by parietal cells further down toward the bottom of the gastric pits. Gastric acid unfolds proteins and activates the stomach enzymes that break the molecular bonds of proteins and lipids. Parietal cells also secrete a chemical named intrinsic factor. Intrinsic factor is necessary for absorption by the intestine of essential vitamin B12
Stretch receptors in the wall of the stomach send afferent information through the vagus nerve to the central nervous system (CNS). The efferent response of the CNS to the sensory information travels back to the gastric pits in the vagus nerve forming a reflex arc within the nerve. Other sensory information including sight, smell and taste also influence vagus nerve efferent activity at parietal cells. The neural reflex arcs match stomach contractions and secretions to the intake of food.
Parietal cell secretion of acid is also increased by Gastrin, a hormone produced by the G-Cells of the gastric pits at the pyloric end of the stomach. I will cover the hormones of the digestive system in another article, but you can read about the basic properties of the endocrine system by clicking here.
Chief cells reside at the bottom of the gastric pits. In response to food in the stomach, chief cells secrete gastric lipase and an inactive precursor of a protein-digesting enzyme called pepsinogen. Pepsinogen is converted to the active enzyme pepsin by the acid environment of the stomach. Gastric lipase is an enzyme that begins the digestion of lipids in the stomach. The activity of gastric lipase is optimal at pH 3-6.
Digestion in the small intestine
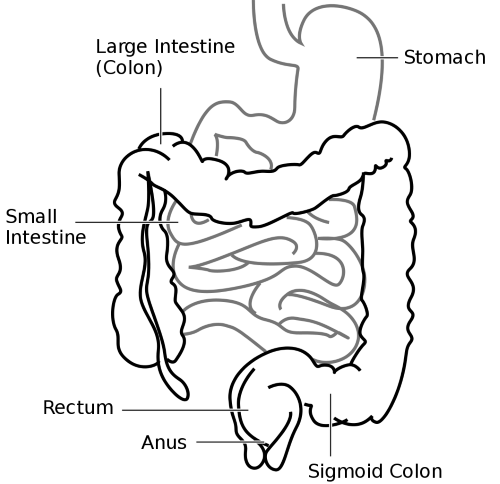
Illustration of the position of the small intestine, Public Domain/Wikimedia Commons
Digestion in the small intestine begins when the stomach releases its partially processed food particle in small portions to the small intestine through its pyloric valve. The small intestine consists of three parts, the duodenum, jejunum and ilium. The duodenum is the section proximal to the stomach, and the ilium is the most distal portion ending at the ascending colon.
Fats, carbohydrate, protein and nucleic acids (RNA and DNA) are broken down into fatty acids, glycerol, monosaccharides, amino acids and nucleotides in the small intestine.
To do this a large surface area is required. The intestinal surface area, the lamina propria is increased by folds upon its folds to form the gastric mucosa. The large first level of folds increases the area of the gastric mucosa by about three-times. Lining the walls of the primary folds are secondary folds named villi. Villi are composed of epithelia cells called enterocytes. The villi increase the surface area another 10-times.
The apical membrane of the enterocyte facing the lumen is covered by microvilli that increase the surface another 20-times. Interspersed among the enterocytes, which are absorptive cells, are goblet cells that secret mucus.
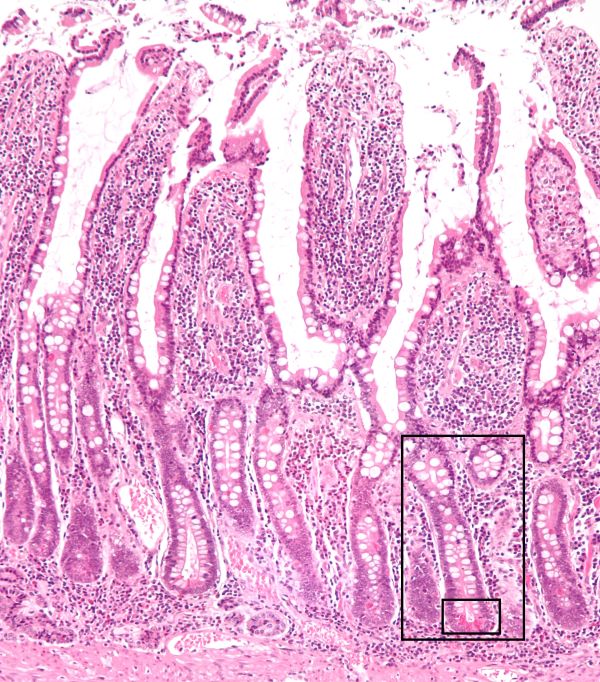
Small intestine villi showing position of the crypts of Lieberkuhn, Nephron/Wikimedia Commons
Between the villi are pits called the crypts of Lieberkϋhn. A crypt in this photomicrograph is outlined by the black boxes. Some crypt cells secrete water and electrolytes obtained from blood.
The crypts also contain stem cells to renew the villus lining of enterocytes. The entire enterocyte epithelium renews every 4-5 days with new cells moving up from the bottom of the villi. The contents of the dying cells that are sluffed off into the lumen of the intestine are thought to contribute to the digestive process.
The nerve supply and the blood and lymph capillaries of the small intestine reside in the center of the villi.
Exocrine pancreas secretions
Distension of the stomach as well as sight, smell and taste of food stimulate exocrine pancreas secretion through vagus nerve efferent activity.
The pancreas delivers a watery solution through its duct to the duodenum. This exocrine secretion contains a variety of enzymes capable of breaking molecular bonds of protein, carbohydrate, lipid and nucleic acids.
Protein digestion
Gastric acid is enough to unfold the tertiary structure of proteins. But proteins are large complex molecules, and it takes a diversity of enzymes to completely break it down to a pool of free amino acids that can be absorbed by blood capillaries. Enzymes are also protein, and most enzymes require a near neutral pH for optimal activity. Bicarbonate in the pancreatic secretion neutralizes gastric acid delivered by the stomach to the small intestine.
Pancreatic proteolytic enzymes arrive at the small intestine in an inactive form. An enzyme called enterokinase secreted by enterocytes of the intestinal villi begins a sequence of chemical steps that activates the proteolytic pancreatic enzymes. Pancreatic enzymes break unfolded proteins into a mixture of free amino acids and small sections of proteins called peptides.
Carbohydrate digestion
Pancreatic amylase for carbohydrate digestion is secreted in its active form. Carbohydrates are sugar molecules also known as saccharides. A single sugar molecule like blood glucose is called a monosaccharide, and long chains of sugar molecules are called oligosaccharides. Humans have enzymes that break some but not all the molecular bonds found in oligosaccharides. Undigestible oligosaccharide material makes up part of dietary fiber. Pancreatic amylase breaks digestible oligosaccharides into strings of 2-4 sugar molecules in the lumen of the duodenum.
Lipid digestion
Lipid digestion first requires overcoming their tendency to cluster together in watery solutions. Emulsification is the dispersion of large lipid clusters into many tiny droplets. The droplets provide a larger surface area for attack by pancreatic lipase. Bile produced in the liver assists in the emulsification process.
Bile is delivered by the common hepatic duct. The common hepatic duct joins the pancreatic duct as it enters the duodenum. An off shoot of the hepatic duct, prior to joining the pancreatic duct fills the gall bladder. The gall bladder provides a secondary source of bile for meals large in fat. In the absence of the gall bladder the liver continuously delivers bile to the small intestine.
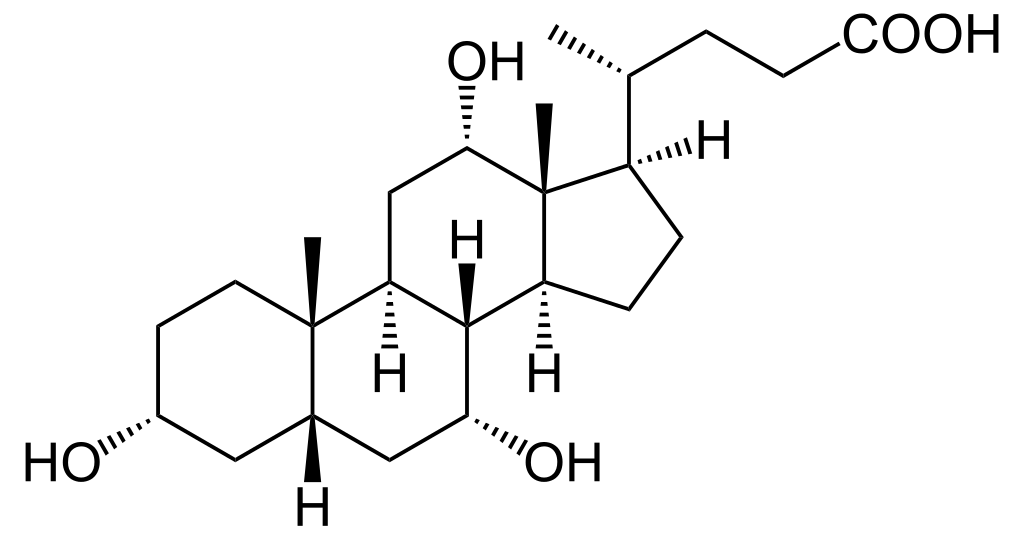
Cholic acid, a bile acid produced by the liver, Public Domain/Wikimedia Commons
Bile is a mixture of bile acids, phospholipids, cholesterol, bile pigments, mucin, electrolytes and the liver’s metabolic waste. It is the bile acids that emulsify the fat droplets. Bile acids are cholesterol derivatives where one end of the molecule at carbon 24 is metabolized to a carboxy acid (-COOH).
When the bile acid molecule is in its normal conformation in solution, the acid end and hydroxyls on three of the rings form a hydrophilic face that dissolves well in water. The rest of the steroid ring structure is itself a lipid that dissolves in the lipid of the droplets.
Lipolytic enzymes in the pancreatic secretion to the small intestine include pancreatic lipase, pancreatic colipase, phospholipase A2 and cholesterol esterase. Pancreatic colipase with the help of bile acids anchor pancreatic lipase to the lipid droplets. All the lipid hydrolysis products are removed from the lipid droplets by forming complexes with bile acids and phospholipids of the bile. These aggregates are called mixed micelles.
Hepatic portal vein
The venous drainage of the small intestine is the hepatic portal vein. The enterocytes of the intestinal villi have multiple transporters on their apical and basal membranes for transferring specific amino acids, monosaccharides nucleotides and lipid hydrolysis products from the lumen of the small intestine to the hepatic portal system and lymphatic capillaries of the villi.
Peptides taken up by enterocytes are reduced to 2-3 amino acid pieces by a group of enzymes located along the enterocyte microvilli. Peptide transporters take the pieces into the enterocyte cell where some are further broken to single amino acids. Other transporters on the basolateral cell membrane, the side of the enterocyte lined by blood vessels, transfer the amino acids out of the cells for pick up by blood vessels at the center of the villi.
Short strings of saccharides produced by luminal amylase are further digested by enterocyte microvilli enzymes to monosaccharides. Monosaccharides are moved into the enterocyte cell by specialized apical membrane transporters. Like amino acids, monosaccharides are moved at the basolateral cell membrane of enterocytes out of the cell into blood vessels by specialized saccharide transporters.
Mixed micelles are taken up at the microvilli into the enterocytes by endocytosis. The endocytosis vesicles merge with the endoplasmic reticulum where the lipid are packages into lipoprotein. Lipoproteins are particles of lipid coated with protein to make the particles soluble in blood. The lipoproteins made by the intestine are named chylomicrons, very low density lipoprotein (VLDL) and high density lipoprotein (HDL).
Protein packaged lipids are secreted at the basolateral membrane of the enterocytes into blood and lymph vessels. The large chylomicron is only picked up by lymph vessels for delivery to blood at the heart. The lymph transports about 50% of absorbed lipids. The remaining 50% are delivered in the venous circulation to the liver’ hepatic portal system as VLDL and HDL.
Large Intestine
The large intestine begins with the ascending colon and ends at the rectum. The movement of undigested material from the terminal end of the small intestine, the ilium, into the cecum of the ascending colon is by bolus movement through the ileocecal sphincter. Bolus movements are separated by periods of no flow. Distension of the colon produces a reflex relaxation of the ileum and contraction of sphincter preventing reflux of colon contents into the ileum.
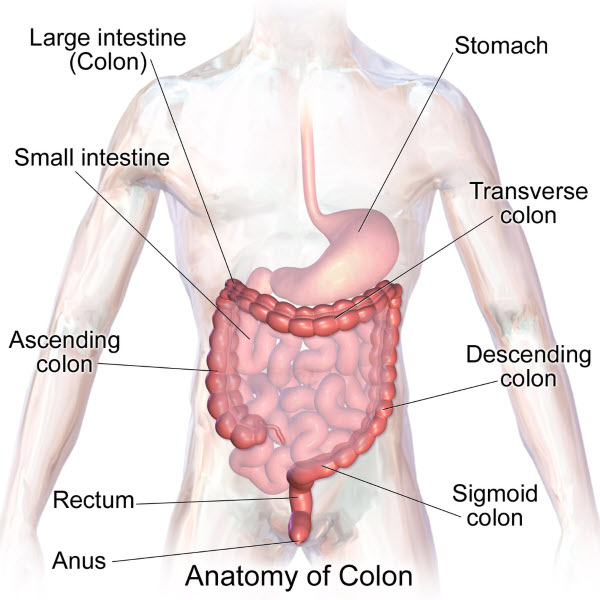
Human colon sections, Bruce Blaus/Shutterstock.com
The cecum of the colon begins at the ileocecal sphincter. From the cecum the parts of the colon are the ascending colon, transverse colon, descending colon, sigmoid colon, rectum and anus. Colon muscle contraction mixes and compacts the undigested material as it moves it from the cecum to the rectum.
A significant part of the water of digestive secretions is reabsorbed into the circulatory system in the small intestine due to osmotic pressure developed as nutrients are transported out of the enterocytes. Remaining water is absorbed into the circulatory system along the length of the colon.
Eating a meal increases muscle activity throughout the length of the colon. This is called the gastrocolic reflex. The gastrocolic reflex can produce mass movement of feces and stimulate defecation. The sensory afferents for this reflex are stretch receptors in the stomach and chemoreceptors in the small intestine.
There are two sphincters controlling release of feces from the rectum. They are the internal anal sphincter under control of the sympathetic and parasympathetic nervous system and the external anal sphincter under voluntary skeletal muscle control.
Digestive system blood flow
Now that you know how important the circulatory system is to digestion, it would be good to review the large blood vessels servicing the digestive system. The arrangement is unusual in that the venous flow from the digestive tract goes to the hepatic portal system of the liver rather than to the inferior vena cava.
The liver is the accessory organ to the digestive system responsible for using molecules acquired in digestion to meet the body’s structural and functional needs. The liver produces blood proteins from amino acids, regulates blood levels of amino acids, converts excess glucose to its storage form glycogen, produces cholesterol and other fats for distribution to other parts of the body as lipoprotein, produces high energy molecules of ATP from glucose and makes glucose as needed between meals from protein and fats.
You can review the arterial and venous blood flow of the digestive System by watching two videos produced by TheAnatomyZone for YouTube. To access them click on their titles below.
Arterial Circulation of the Digestive System
Venous Drainage of the Digestive System
Further reading
Gastrointestinal Tract Neural Regulation
Gastrointestinal Hormones for Digestion & Fasting
3 Simple Secrets to Learning Physiology
Do you have questions?
Send me an email with your questions at DrReece@MedicalScienceNavigator.com or put them in the comment box. I always read my mail and answer it. Please share this article with your fellow students taking anatomy and physiology by clicking your favorite social media button.

Margaret Thompson Reece PhD, physiologist, former Senior Scientist and Laboratory Director at academic medical centers in California, New York and Massachusetts is now Manager at Reece Biomedical Consulting LLC.
She taught physiology for over 30 years to undergraduate and graduate students, at two- and four-year colleges, in the classroom and in the research laboratory. Her books “Physiology: Custom-Designed Chemistry”, “Inside the Closed World of the Brain”, and her online course “30-Day Challenge: Craft Your Plan for Learning Physiology”, and “Busy Student’s Anatomy & Physiology Study Journal” are created for those planning a career in healthcare. More about her books is available at https://www.amazon.com/author/margaretreece. You may contact Dr. Reece at DrReece@MedicalScienceNavigator.com, or on LinkedIn.
Dr. Reece offers a free 30 minute “how-to-get-started” phone conference to students struggling with human anatomy and physiology. Schedule an appointment by email at DrReece@MedicalScienceNavigator.com.
Comments
Human Digestion: Main & Accessory Organs — No Comments
HTML tags allowed in your comment: <a href="" title=""> <abbr title=""> <acronym title=""> <b> <blockquote cite=""> <cite> <code> <del datetime=""> <em> <i> <q cite=""> <s> <strike> <strong>